Great Tit

Introduction
The Great Tit is a common visitor to garden bird feeders. Outside urban areas, it is primarily a species of scrub and deciduous woodland.
The Great Tit is a striking little bird, with a bluish-green back and a yellow breast divided by a central black stripe. The sexes are similar, but in males the black stripe is more pronounced. It is a vocal species with a huge range of songs and calls; its most familiar is its repeated 'teacher-teacher-teacher' song which can be heard throughout woodlands in spring.
UK Great Tit numbers increased from the 1960s, although they have stablised since about 2010. Great Tits take readily to nest boxes, and lay one (sometimes two) clutches a year. They are primarily insectivorous in summer, but over winter they frequent bird feeders and are often found in mixed flocks with other tit species. They are widespread throughout Britain & Ireland apart from on some islands in northern and western Scotland.
- Our Trends Explorer gives you the latest insight into how this species' population is changing.
Key Stats
Identification
ID Videos
This section features BTO training videos headlining this species, or featuring it as a potential confusion species.
GBW: Blue Tit and Great Tit
#BirdSongBasics: Great Tit and Coal Tit
#BirdSongBasics: Blue Tit and Great Tit
Songs and Calls
Song:
Call:
Begging call:
Status and Trends
Conservation Status
Population Change
Great Tit numbers have increased fairly steadily since the 1960s, with the exception of two or three brief periods of stability or shallow decline. The BBS map of change in relative density between 1994-96 and 2007-09 indicates that increase during that period occurred throughout the UK range. A new strain of avian pox, first recorded in 2006, is increasing in Great Tits but without evidence so far of demographic effects (Lawson et al. 2012a, 2018). Laying dates have advanced by nine days since 1968. Earlier nesting can be linked to the phenology of spring greening in woodland, though with much small-scale spatial variation (Cole et al. 2015). There has been an increase across Europe since 1980 (PECBMS: PECBMS 2020a>).
Distribution
Great Tits occur in 90% of 10-km squares in Britain & Ireland in both winter and the breeding season. As with the Blue Tit, the only significant range gaps are in treeless moorland areas of the Scottish Highlands, the Outer Hebrides and Northern Isles. Densities are highest in lowland wooded landscapes of England, Wales and parts of eastern Ireland.
Occupied 10-km squares in UK
2007/08–10/11
or view it on Bird Atlas Mapstore.
2008–11
or view it on Bird Atlas Mapstore.
European Distribution Map
Distribution Change
Change in range size has been small over the last 40 years, but in many parts of the range, and especially in Ireland, there have been large increases in abundance.
Change in occupied 10-km squares in the UK
from 1981–84 to 2007–11
or view it on Bird Atlas Mapstore.
from 1968–72 to 2008–11
or view it on Bird Atlas Mapstore.
Seasonality
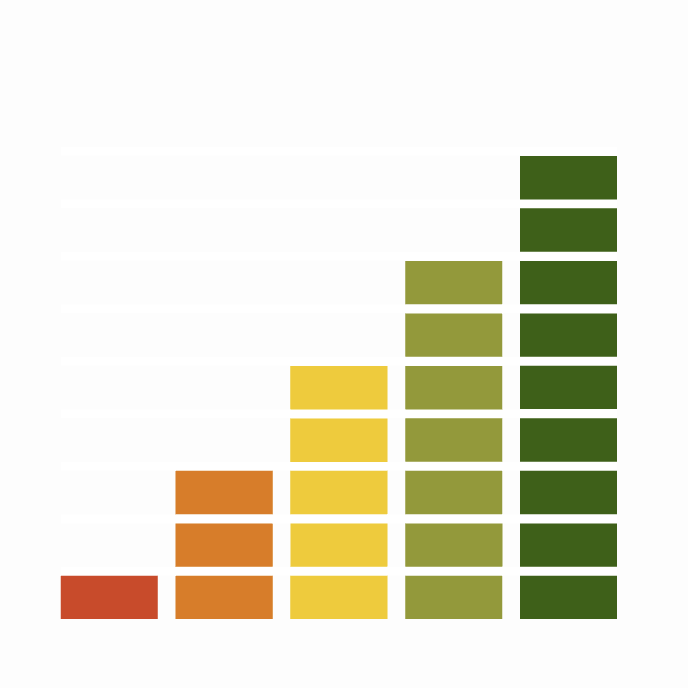
Great Tit is recorded throughout the year on up to 60% of complete lists.
Weekly pattern of occurrence
The graph shows when the species is present in the UK, with taller bars indicating a higher likelihood of encountering the species in appropriate regions and habitats.

Habitats
Breeding season habitats
Relative frequency by habitat
The graph shows the habitats occupied in the breeding season, with the most utilised habitats shown at the top. Bars of similar size indicate the species is equally likely to be recorded in those habitats.

Movement
Britain & Ireland movement
Foreign locations of birds ringed or recovered in Britain & Ireland
Dots show the foreign destinations of birds ringed in Britain & Ireland, and the origins of birds ringed overseas that were subsequently recaptured, resighted or found dead in Britain & Ireland. Dot colours indicate the time of year that the species was present at the location.
- Winter (Nov-Feb)
- Spring (Mar-Apr)
- Summer (May-Jul)
- Autumn (Aug-Oct)

European movements
EuroBirdPortal uses birdwatcher's records, such as those logged in BirdTrack to map the flows of birds as they arrive and depart Europe. See maps for this species here.
The Eurasian-African Migration Atlas shows movements of individual birds ringed or recovered in Europe. See maps for this species here.
Biology
Productivity and Nesting
Nesting timing
Egg measurements
Clutch Size
Incubation
Fledging
Survival and Longevity
Survival is shown as the proportion of birds surviving from one year to the next and is derived from bird ringing data. It can also be used to estimate how long birds typically live.
View number ringed each year in the Online Ringing Report.
Lifespan
Survival of adults
Survival of juveniles
Biometrics
Wing length and body weights are from live birds (source).
Wing length
Body weight
Ring Size
Classification, names and codes
Classification and Codes
- Order: Passeriformes
- Family: Paridae
- Scientific name: Parus major
- Authority: Linnaeus, 1758
- BTO 2-letter code: GT
- BTO 5-letter code: GRETI
- Euring code number: 14640
Alternate species names
- Catalan: mallerenga carbonera
- Czech: sýkora konadra
- Danish: Musvit
- Dutch: Koolmees
- Estonian: rasvatihane
- Finnish: talitiainen
- French: Mésange charbonnière
- Gaelic: Currac-baintighearna
- German: Kohlmeise
- Hungarian: széncinege
- Icelandic: Flotmeisa
- Irish: Meantán Mór
- Italian: Cinciallegra
- Latvian: liela zilite
- Lithuanian: didžioji zyle
- Norwegian: Kjøttmeis
- Polish: bogatka (zwyczajna)
- Portuguese: chapim-real
- Slovak: sýkorka velká
- Slovenian: velika sinica
- Spanish: Carbonero común
- Swedish: talgoxe
- Welsh: Titw Mawr
- English folkname(s): Titmouse
Research
Causes of Change and Solutions
Causes of change
Demographic trends in breeding parameters do not suggest that increases in this species are due to improvements in breeding performance. There is some evidence, albeit limited, that improvements in survival rates, due to amelioration in wintering conditions, may have been responsible. Evidence for ecological drivers of the population increase is limited but increased provisioning in gardens and milder winters may have played a role.
Further information on causes of change
The number of fledglings per breeding attempt have decreased alongside decreases in clutch and brood sizes (see above). Daily failure rates at the egg stage have also decreased but daily failure rates at the chick stage has not changed. Consequently, breeding success does not contribute substantially to population change, and integrated modelling confirms that variation in adult survival is the primary driver of annual population change (Robinson et al. 2014).
Increases in survival rates, due to more widespread food provision in gardens during winter is one possible explanation for the increase (Plummer et al. 2019). Horak & Lebreton (1998) found that survival rates in Estonia were higher in urban populations than rural ones and suggested that this was partly due to supplementary feeding in gardens. Increasing winter temperature may have also played a role. Ahola et al. (2009) suggested that, for their study population in Sweden, increasingly favourable conditions in winters have enhanced the survival rates of Great Tit and resulted in the observed increase in Great Tit breeding density.
Other factors are also likely to influence survival rates. There is some evidence that the beech crop may be influential and it has been shown that survival rates can be related to beechmast production (Verhulst 1992, Perdeck et al. 2000), although there is no evidence that beechmast production has gone up. Perdeck et al. (2000) provided further evidence for this as supplemental food increased survival of both juveniles and adults, supporting the winter-food limitation hypothesis. In a Finnish population, Orell (1989) reported that the high survival rates of resident juveniles after a warm August may be attributable to food availability during the time when the birds undergo their post-juvenile moult.
Great Tits have advanced their laying date, in line with climatic change. This has been found by several studies (e.g. Sanz 2002, Visser et al. 2009, Bauer et al. 2010), but does not seem to be influencing the population trend. Intraspecific competition may also drive population changes in Great tits (Gamelon et al. 2019), and there is evidence that density-dependent effects influence clutch size, hatching success and fledging success, which may explain the decreases observed in clutch size as fledging rates as the population has increased (Bodey et al. 2020). Severe weather events are expected to become more frequent as a result of climate change and conditions such as heavy and persistent rainfall can also be an important driver of Great Tit breeding success (Scholl et al. 2020); however, there is no evidence that this has driven population change.
Information about conservation actions
Like its close relative the Blue Tit, the population of this species has increased since the 1970s, hence it is not a species of concern and no conservation actions are currently required. However, it has experienced a shallow decline over the last ten years and therefore ongoing monitoring would be prudent.
Ongoing provision of garden bird food is likely to continue to benefit the Great Tit. However, the effects are not always entirely positive and feeders may contribute to the spread of avian pox, so those providing food should ensure they follow good hygiene practices. The provision of nest boxes, both in gardens and elsewhere, is also likely to continue to benefit this species.
Publications (8)
Declines in invertebrates and birds – could they be linked by climate change?
Author:
Published: 2023
The long-term declines evident in many bird and invertebrate species have their origins within a suite of potential drivers, one of which is climate change. As well as impacting bird species directly, could climate change be increasingly hitting bird populations through its impacts on their invertebrate prey?
09.01.23
Papers

Temporal avoidance as a means of reducing competition between sympatric species
Author:
Published: 2023
Human activities modify the availability of natural resources for other species, including birds, and may alter the relationships between them. The provision of supplementary food at garden feeding stations, for example, might favour some species over others and change the competitive balance between them. This paper investigates the behavioural responses to competition of the Marsh Tit, a species that is subordinate to both the Blue Tit and the Great Tit.
24.05.23
Papers

A review of the impacts of air pollution on terrestrial birds
Author:
Published: 2023
A review paper by BTO considers 203 studies of the effects of air pollution on 231 bird species. Of these studies, 82% document at least one negative effect associated with increasing levels of pollution. The review also highlights biases towards particular study species, especially Great Tit and Pied Flycatcher, and also towards particular geographical regions (Western Europe) and pollutants (heavy metals). The paper proposes research approaches that could help to provide a fuller understanding of how birds are impacted by air pollution.
15.05.23
Papers

Evidence that rural wintering populations supplement suburban breeding populations
Author:
Published: 2022
Urban areas can and do hold significant populations of birds, but we know surprisingly little about how these populations are connected with those present within the wider countryside. It has been suggested that the populations using these different habitats may be linked through seasonal movements, with individuals breeding in rural areas moving into urban sites during the winter months to exploit the supplementary food provided at garden feeding stations. However, little work has been done to test this hypothesis.
24.11.22
Papers Bird Study
The effects of a decade of agri-environment intervention in a lowland farm landscape on population trends of birds and butterflies
Author:
Published: 2022
Food production and wildlife conservation are often thought of as incompatible goals, and it is rare that conservation studies consider both economics and long-term changes in ecology. However, a decade-long study at a commercial arable farm in Buckinghamshire has found that agri-environment schemes significantly increased local bird and butterfly populations without damaging food production, offering hope for the UK’s farmland birds and butterflies.
01.08.22
Papers

Tritrophic phenological match-mismatch in space and time
Author:
Published: Spring 2018
The increasing temperatures associated with a changing climate may disrupt ecological systems, including by affecting the timing of key events. If events within different trophic levels are affected in different ways then this can lead to what is known as phenological mismatch. But what is the evidence for trophic mismatch, and are there spatial or temporal patterns within the UK that might point to mismatch as a driver of regional declines in key insect-eating birds? A changing climate is leading to changes in the timing of key ecological events, including the timing of bud burst, the spring peak in leaf-eating caterpillar biomass and the timing of egg-laying in many bird species. If the timings of these different events shift at different rates then there is a danger that they may get out of synch with one another, something that is referred to as phenological mismatch. This may be a particular problem for birds like Blue Tit, Great Tit and Pied Flycatcher, which time their breeding attempts to exploit the spring peak in caterpillar abundance. Much of the recent work on mismatch and its impacts on the fitness and population trends of caterpillar-eating birds has looked at changes over time. However, it is also possible for mismatch to vary in space if species respond differently in different areas, perhaps because of local adaptation to geographic variation in the cues that they use. This paper looks at mismatch in both space and time, using information from three trophic levels, namely trees, caterpillars and caterpillar-eating birds. While information on bud burst came from 10,000 observations of oak first leafing for the period 1998-2016, that for caterpillar biomass was inferred from frass traps set beneath oak trees at sites across the UK for the period 2008-2016. Bird phenology data came from the ‘first egg date’ values calculated from 85,000 nest records of Blue Tit, Great Tit and Pied Flycatcher. The focus of the work was on the relationship between the phenologies of these interacting species; where timing changes more in one species than the other, this is indicative of spatial or temporal variation in the magnitude of mismatch. The results reveal that, for the average latitude (52.63°N) and year, there is a 27.6 day interval between the timing of oak first leaf and peak caterpillar biomass. With increasing latitude, the delay in oak leafing is significantly steeper than that of the caterpillar peak. At 56°N the predicted interval between these two trophic levels drops to 22 days. In the average year and at the average latitude, the first egg dates of Blue Tits and Great Tits were roughly a month earlier than peak caterpillar biomass, meaning that peak demand for hungry chicks occurred soon after the peak in resource availability. Interestingly, peak demand in Pied Flycatchers occurred nearly two weeks later than peak caterpillar availability, suggesting a substantial trophic mismatch between demand and availability for this species within the UK. However, it is worth noting that Pied Flycatchers provision their nestlings with fewer caterpillars and more winged invertebrates compared to the tit species studied, so they may be less dependent on the caterpillar peaks. The work also revealed that the timing of first egg date between years varied by less than the variation seen in timing of the caterpillar resource peak, which gave rise to year-to-year variation in the degree of mismatch. For every 10 day advance in the caterpillar peak, the corresponding advance in the three bird species is 5.0 days (Blue Tit), 5.3 days (Great Tit) and 3.4 days (Pied Flycatcher). In late springs, peak demand from the tits is expected to coincide with the peak resource availability, with flycatcher demand occurring shortly after. In early springs, the peak demand of nestlings of all three species falls substantially later than the peak, leaving the three mismatched. Warmer conditions also shortened the duration of caterpillar peaks. One of the key findings of the work is that in the average year there is little latitudinal variation in the degree of caterpillar-bird mismatch. This means that more negative declines in population trends of certain insectivorous birds in the southern UK, driven by productivity, are unlikely to have been driven by greater mismatch in the south than the north. The lack of evidence for latitudinal variation in mismatch between these bird species and their caterpillar prey suggests that mismatch is unlikely to be the driver of the spatially varying population trends found in these and related species within the UK.
23.04.18
Papers

Health hazards to wild birds and risk factors associated with anthropogenic food provisioning
Author:
Published: 2018
Work on emerging infectious diseases and garden birds in the UK has been supported by citizen science projects, most notably Garden BirdWatch, Garden Wildlife Health and the Garden Bird Health Initiative – the latter now superseded by Garden Wildlife Health. Through these schemes, researchers have been able to carry out national surveillance of emerging diseases, including finch trichomonosis, Paridae pox and passerine salmonellosis. This paper, part of a special issue of Philosophical Transactions focusing on wildlife disease issues, reviews the work that has been carried out on these diseases over the past 25 years. It also takes a look at the occurrence of mycotoxin contamination of food residues in bird feeders, which also pose a risk to the health of wild birds.
12.03.18
Papers
Passerines may be sufficiently plastic to track temperature-mediated shifts in optimum lay date
Author:
Published: 2016
19.05.16
Papers
More Evidence
More evidence from Conservation Evidence.com
Partners



Citing BirdFacts
If you wish to cite particular content in this page (e.g. a specific value) it is best to use the original sources as linked in the page. For a more general citation of the whole page please use: BTO (20XX) BirdFacts Species: profiles of birds occurring in the United Kingdom. BTO, Thetford (www.bto.org/birdfacts, accessed on xx/xx/xxxx).